Taylor-Clark Lab
Research
Overview
Every organ is innervated by a heterogeneous population of sensory nerves that detect specific stimuli (e.g. mechanical pressure, temperature, pH, inflammatory mediators) due to the expression of receptors and ion channels. Sensory nerves conduct this information to the CNS and, depending on the nerve subtype and originating organ, evoke specific behavioral, emotional and reflex responses. Our main focus is on nociceptive sensory nerves within the lungs and airways. These nerves are activated by inflammation (e.g. asthma), infections and inhaled pollutants and irritants and cause cough, wheeze, decreased airflow and mucus secretion, as well as modulating cardiovascular function.
-
Mitochondria signaling
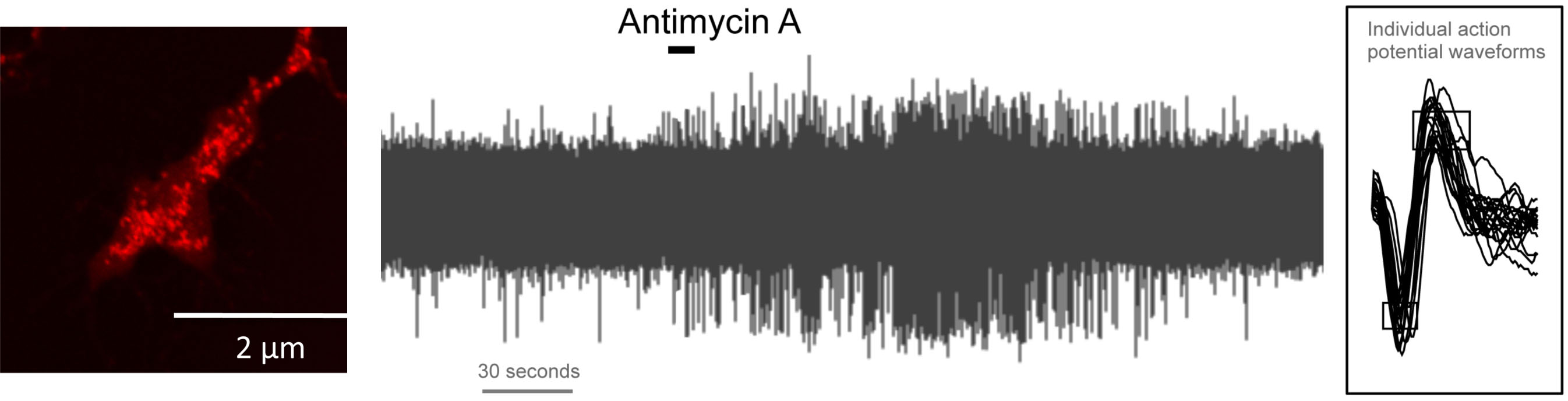
Sensory nerve terminals are packed with mitochondria. Multiple environmental stimuli, including inflammation, regulate mitochondrial function. Mitochondria modulate cellular signaling via production of ATP and reactive oxygen species (ROS) and the regulation of Ca2+ fluxes. We have found that mitochondrial signaling selectively activates nociceptive sensory nerves. These responses were dependent on the nociceptive ion channels transient receptor potential (TRP) ankyrin 1 (A1) and TRP vanilloid 1 (V1). Furthermore, mitochondrial signaling also independently caused an increase in nociceptive sensory nerve excitability. This hyperexcitability was not dependent on TRPA1 or TRPV1 but instead was due to the activity of protein kinase C.
Mitochondria (red) in sensory terminal in vitro; initiation of action potential discharge in vagal sensory nociceptor with mitochondrial modulation by antimycin A. -
TRPA1
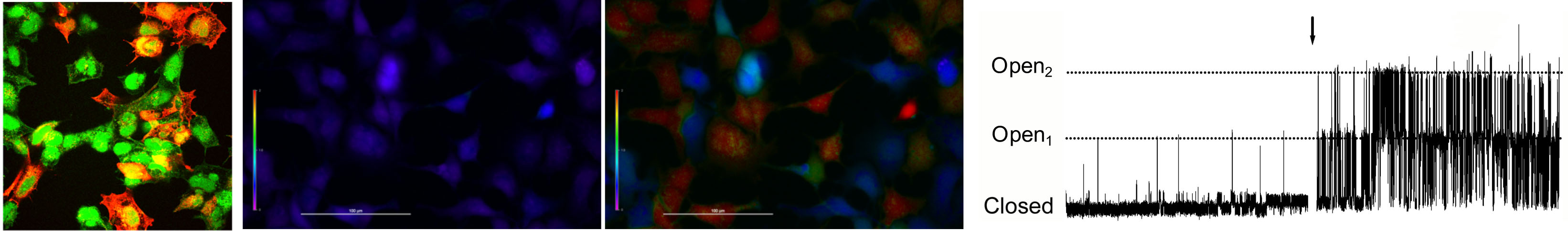
Transient receptor potential ankyrin 1 (TRPA1) is a polymodal cation channel expressed on a subset of nociceptive sensory nerves. Best known as the ‘wasabi detector’, TRPA1 is activated by a host of environmental and inflammatory stimuli. We have previously characterized TRPA1 as the molecular mechanism underlying nociceptor activation by the environmental pollutants ozone and toluene diisocyanate and numerous products of inflammation and oxidative stress including 4-hydroxynonenal, prostanoids and 9-nitrooleate. These molecules activate TRPA1 via covalent modification of specific cysteine residues on the channel. We have recently shown that these particular cysteines demonstrate exceptionally high reactivity, thus accounting for their role in noxious stimuli detection.
From left: TRPA1 (red) expressed in HEK293 cells (green); low cytosolic calcium (blue) in TRPA1-expressing HEK293 in control conditions; high cytosolic calcium (red) in TRPA1-expressing HEK293 treated with wasabi; single TRPA1 channel currents in TRPA1-expressing HEK293 in control conditions and with wasabi treatment. -
Sensory nerve innervation
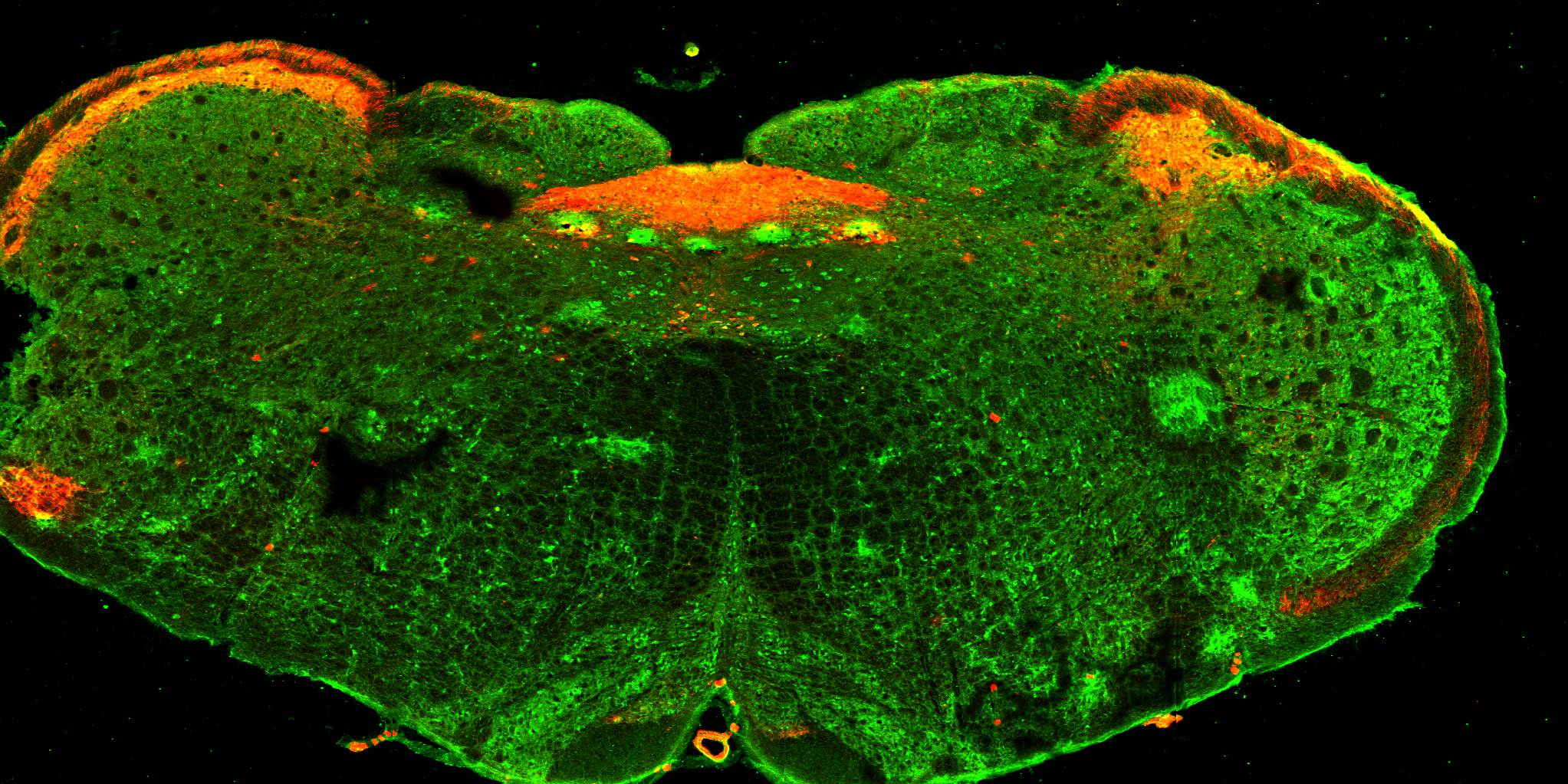
Fundamentally, sensory nerve function can be most generally described by three parameters: what the nerve detects; where in the body is the sensor; and what CNS circuits are regulated by the nerve activity? Using state-of-the-art genetic tools, we are investigating the specific neural pathways that connect specific sensory subsets to the CNS. In this way we are mapping the sensory arm of the peripheral nervous system.
Brainstem showing the location of nociceptive sensory nerve fibers (red) -
Pulmonary-cardiac reflexes
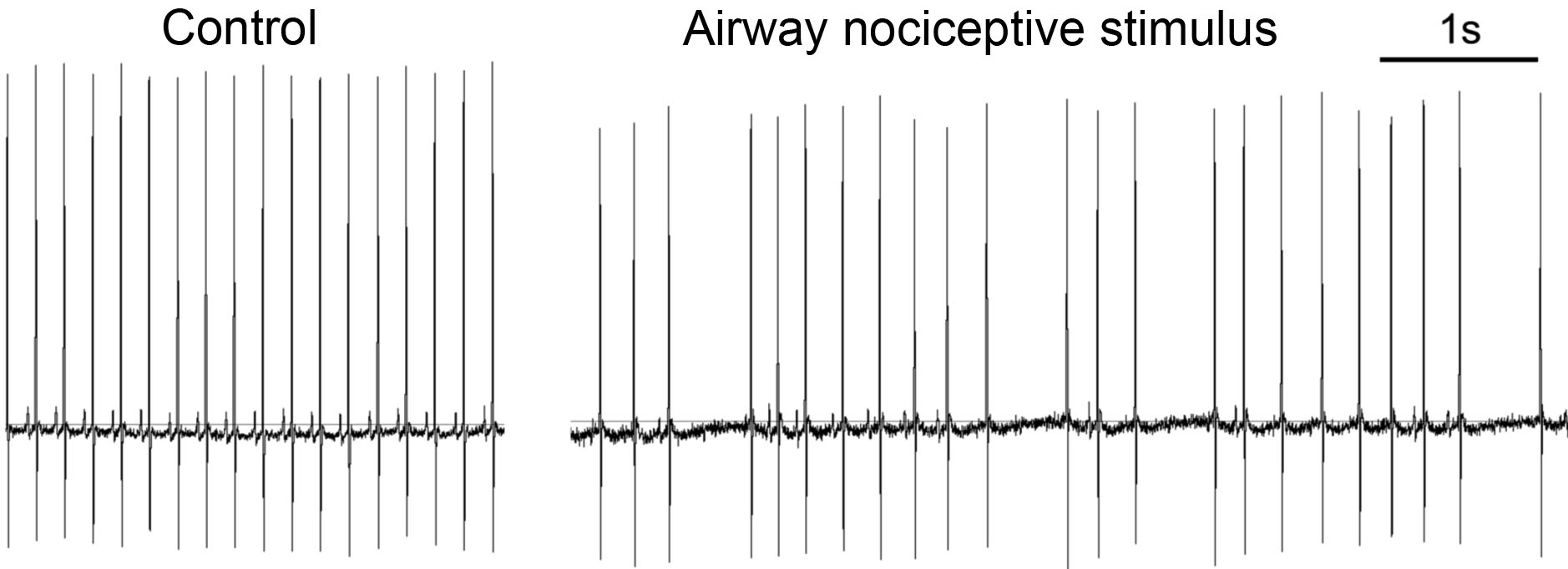
Oxygen consumption by working tissues (e.g. brain, muscles) is linked with blood flow. As such the CNS integrates the control of the respiratory and cardiovascular physiology. Part of this integration are reflexes initiated from the airways which can impact cardiovascular function. This may be harmless in healthy individuals, but represents a major risk factor for individuals with preexisting cardiovascular disease. We have recently shown that inhalation of nociceptive stimuli cause chaotic slowing of heart rhythms (bradycardia) due to reflex stimulation of cardiac parasympathetic nerves downstream of airway nociceptive signaling. We are interested in understanding how preexisting cardiovascular function modulates these pulmonary-cardiac reflexes.
Electrocardiogram at rest and with airway nociceptive stimulus
Selected Publications
Taylor-Clark TE, Undem BJ, MacGlashan DW Jr, Ghatta S, Carr MJ, McAlexander MA. Prostaglandin-induced activation of nociceptive neurons via direct interaction with transient receptor potential A1 (TRPA1). Mol Pharmacol. 2008 Feb;73(2):274-81.
Taylor-Clark TE, McAlexander MA, Nassenstein C, Sheardown SA, Wilson S, Thornton J, Carr MJ, Undem BJ. Relative contributions of TRPA1 and TRPV1 channels in the activation of vagal bronchopulmonary C-fibers by the endogenous autacoid 4-oxononenal. J Physiol. 2008 Jul; 586(14): 3447-59.
Taylor-Clark TE, Undem BJ. “Ozone activates airway nerves via the selective stimulation of TRPA1 ion channels”, J Physiol. 2010 Feb; 588(3): 423-33.
Nesuashvili L, Hadley SH, Bahia PK, Taylor-Clark TE. Sensory nerve terminal mitochondrial dysfunction activates airway sensory nerves via transient receptor potential (TRP) channels. Mol Pharmacol. 2013 May;83(5):1007-19. PMID: 23444014, PMCID: 3629826.
Hadley SH, Bahia PK, Taylor-Clark TE. Sensory Nerve Terminal Mitochondrial Dysfunction Induces Hyperexcitability in Airway Nociceptors Via Protein Kinase C. Mol Pharmacol. 2014 June;85(6):839-48. PMID: 24642367, PMCID: 4014670.
Hooper JS, Hadley SH, Morris KF, Breslin JW, Dean JB, Taylor-Clark TE. Characterization of cardiovascular reflexes evoked by airway stimulation with allylisothiocyanate, capsaicin and ATP in Sprague-Dawley rats. J Appl Physiol (1985). 2016; 120(6):580-91.
Bahia PK, Parks TP, Stanford KR, Mitchell DA, Varma S, Stevens Jr. SM, Taylor-Clark TE. The exceptionally high reactivity of Cys 621 is critical for electrophilic activation of the sensory nerve ion channel TRPA1. J Gen Physiol. 2016 Jun;147(6):451-65.